Восточноафриканский гепард
Acinonyx jubatus raineyi (Heller, 1913)

Восточно-африканский или танзанийский гепард, реже кенийский.
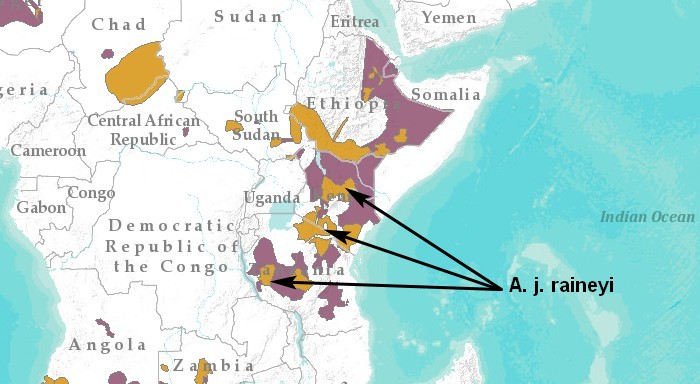
Ареал: Восточная Африка (Кения, юг Сомали, Танзания и Уганда, кроме крайних северных частей Кении и Уганды, подвидовое положение которых спорно). Южная граница ареала подвида указывалась до северного Мозамбика (Heller & Roosevelt, 1914), но сейчас в этой части страны гепард вымер, но чаще гепарды южной Танзании и северного Мозамбика относили к подвиду A. j. junatus.
Возможно исчезнувшим, согласно данным IUCN (2022) вид считается на юге Сомали, окончательно вымершим - в Бурунди и Руанде.
На данный момент в подвид включены обитавшие рядом A. j. ngorongorensis (Танзания от кратера Нгоронгоро до ДР Конго) и A. j. velox (саванны Кении долины Лойта в провинции Рифт-Валли и примыкающие саванны Танзании).
Несколько раз подвид пытались объединить с южно-африканским (A. j. jubatus) на основе генетических исследований. Поэтому в разных источниках подвид считается либо самостоятельным, либо отдельным. Казалось бы, что точку поставила довольно спорная последняя ревизия таксономии кошачьих (Kitchener et al., 2017), однако последние генетические исследования (Prost et al., 2022) в очередной раз подтвердили полную несостоятельность ревизии 2017 г.

Вторая по величине популяции после самой многочисленной южно-африканского гепарда (A. j. jubatus). Согласно IUCN (2022), популяция оценивается в 1569 половозрелых особей, распределенных по 5 субпопуляциям в порядке убывания предполагаемой численности: 1250 половозрелых особей в ландшафте Серенгети / Мара / Цаво / Лайкипиа в Кении и северной Танзании; 184 в ландшафте Руаха в центральной Танзании; 55 в ландшафте Катави-Угалла в Танзании; 47 в степи Масаи в Танзании; 33 в Южной Туркане в Кении.
Значительная часть (75,4%) ареала гепардов в Восточной Африке находится за пределами охраняемых территорий, на землях, которые в основном заняты традиционными скотоводческими общинами, где гепарды сталкиваются с повышенными угрозами со стороны ответных убийств; неустойчивое управление пастбищами, ведущее к потере потенциальной добычи; и минимальная защита от других незаконных убийств (например, в целях торговли).
Гепарды истреблены на большей части территории Танзании и Сомали всего за последние несколько десятилетий. Вполне вероятно, что популяция гепардов во многих странах сталкивается с аналогичным резким сокращением из-за быстрого изменения землепользования и растущей нетерпимости к гепардам за пределами ООПТ, а также массовой торговли мясом диких животных, которая привела к удалению добычи с больших территорий ареал гепарда.

Эта раса представляет собой светлую короткошерстную форму. Основной цвет от охристого до светло-розовато-желтого, наиболее темный по средней линии, где он охристо-коричневатый. Тело равномерно помечено круглыми черными пятнами диаметром ок. 2 см, между которыми иногда встречаются небольшие пятна или точки. Пятна на ногах и животе имеют вытянутую форму и покрывают конечности до пальцев ног. Передние лапы гораздо более заметно пятнистые, чем задние, на которых пятна маленькие или нечеткие возле пальцев. Верх головы и затылок отмечены многочисленными мелкими черными пятнами, а морда охристая без пятен и резко очерчена по бокам черной "слезной" полосой, идущей от глаза ко рту за вибриссами ("усами") и затем проходит назад к углу рта. Бока головы за глазом и под ухом отмечены несколькими черными пятнами, но при этом не образуется четкой линии между глазом и основанием уха. Уши розовато-охристые, с широкой черной полосой, покрывающей нижнюю половину задней стороны. Подбородок и верхняя часть горла от белых до кремовых, живот от кремового до кремово-желтого цвета, с удлиненными пятнами. Шерсть короткая на дорсальной поверхности, ее длина на крупе составляет 2,2 см. Затылок покрыт короткой гривой шириной 7,5 см, идущей от головы до плеч. Средняя линия груди и живота, а также вся длина вентральной поверхности хвоста дополнительно снабжены ершем или короткой гривой из более длинных пушистых волос. Хвост пятнистый, как и спина, с пятью черными кольцами на конце и белым кончиком.

Так как ныне в подвид включены бывшие velox и ngorongorensis, стоит привести и их описания.
Heller (1913), описывая подвид velox, указывал его как африканский высокогорный гепард A. j. velox. Согласно первому описанию, подвид отличался крупными, близко расположенными черными пятнами, которые преобладали над охристым оттенком основного цвета, заметно пятнистыми задними лапами, длинной шерстью и большими размерами тела. От jubatus из Капского региона Африки отличался более крупными пятнами на спине, более светлому основному цвету и большему размеру тела. От raineyi отличался отсутствием розоватого налета на шерсти, более крупным и многочисленным пятнам и более длинной шерстью.
Основной цвет живого первого описанного как ngorongorensis экземпляра из зоопарка Лейпцига, но привезенного из Нгоронгоро (Hilzheimer, 1913), был пергаментно желто-коричневым; снизу очень окраска светлая бледно-серо-желтая, совсем без белого; пятнистость нижней части шеи очень выраженная; щеки сероватые, сильно пятнистые; задняя часть уха желтая, с тонкой черной полоской у основания; подбородок и губы белые; грудь и живот без пятен; внешняя сторона ног до самых пальцев сильно пятнистая; хвост с тремя полными кольцами, кончик желтовато-белый.

Восточно-африканский гепард - крупнейший из подвидов. Первый описанный как raineyi взрослый самец (ныне округ Мачакос, бывшая Восточная провинция, Кения) имел следующие размеры тела: тело с головой 124,5 см, хвост 77, 5 см, задняя стопа 30,5 см, ухо 7,6 см. Здесь интересно заметить, что эти данные привели Хеллер и Рузвельт (1914), а ранее Хеллер (1913) указывал, что измерений не имеется. Взрослый самец с равнин Лойта, описанный как velox, имел более крупные размеры: тело с головой 130 см; хвост 74 см, задняя стопа 30 см; ухо 8 см.
Согласно данным Caro (2013) в 1994 г. в НП Серенгети самцы имели в среднем длину тела 122,5 см (113-136 см), длину хвоста 68 см (63-74 см), вес 41,4 кг (28,5-51,0 кг); самки длину тела 124,5 см (113-140 см), длину хвоста 66 см (60-73), вес 35,9 кг (21-43 кг). В Кении самец и самка, соответственно, имели длину тела с головой 132 и 118 см, длину хвоста 66 и 73 см, вес 61 кг (58–65) и 52 кг (41–63).

Обитает прежде всего на открытых равнинах, лугах, в саваннах, предпочитая засушливые регионы, пригодные для охоты. Встречается также на берегах озер и в прибрежных районах. Однако имеются сообщения о гепардах на высоте 4000 м на горе Кения.
По сравнению с другими крупными кошками плотность гепардов относительно низкая. Самая высокая плотность, не считая небольших и тщательно управляемых огороженных заповедников в Южной Африке, наблюдается в НП Серенгети, где плотность колеблется до 2,5 ос./100 км², но гепарды могут собираться сезонно с повышенной плотностью, что может создавать ошибочное впечатление об общей плотности. Относительно низкая плотность гепардов в богатой травоядными животными Серенгети объясняется межвидовой конкуренцией (особенно с более крупными видами, такими как львы и пятнистые гиены, которые могут убивать детенышей гепарда). Очевидно, что гепарды могут сосуществовать вместе с другими конкурентами и разработали стратегии избегания, чтобы минимизировать потери добычи и детенышей там, где плотность конкурентов высока.

Добыча состоит в основном из копытных мелких и средних размеров, обычно весом менее 40 кг; однако масса добычи варьируется в широком диапазоне: от телят африканского буйвола (Syncerus caffer) и жирафов (Giraffa camelopardalis) до наземных птиц и мелких млекопитающих, в том числе зайцев, дикобразов, крыс и цесарок. Гепарды предпочитают убивать наиболее доступную добычу, присутствующую на территории, в диапазоне массы тела 23–56 кг, в среднем 36 кг. Импала (Aepyceros melampus), газель Томсона (Eudorcas thomsonii) и газель Гранта (Nanger granti) являются наиболее предпочтительными. Предпочитаемые виды добычи обеспечивают минимальный риск травм, а их небольшой размер означает, что гепарды могут съесть их до клептопаразитов.
Разумеется, предпочтения в рационе различаются географически. В Серенгети (Танзания) газели Томсона были наиболее предпочтительной добычей, тогда как в НП Найроби (Кения) тремя излюбленными добычами были импалы (27% из 183 убийств), Гранта Газели (25%) и газели Томсона (22%). В Танзании основной добычей гепарда является газель Томсона на равнинах (Серенгети), но импала в лесных массивах. В засушливых кустарниках северной Кении в качестве основных жертв - куду (Tragelaphus strepsiceros), геренук (Litocranius walleri) и дикдик (Madoqua kirkii).
Кроме вышеуказанных в качестве добычи указываются как травоядные - редунка (Redunca redunca), водяной козел (Kobus ellipsiprymnus), так и мелкие хищники - шакалы (Canis spp. и Lupulella spp.), лисицы (Vulpes spp.) и др.

Во всем ареале предпочтение отдается молодым животным, а не взрослым особям, особенно в случае крупных видов, таких как антилопы гну (Connochaetes taurinus). Например, в НП Серенгети детеныши газелей Томсона составляли 53,7% добычи гепардов, но только 5,9% популяции газелей.
Кроме того, гепарды выбирают копытных, которые находятся поодиночке или небольшими группами, в высокой растительности, менее бдительны, на краю стада и вдали от ближайших соседей. Это часто самцы, хотя гепарды более успешны в поимке одиночных самок газелей Томсона, чем одиночных самцов. Самцы гепардов ловят более крупную добычу, чем самки, из-за своего большего размера и охоты коалициями. Однако взрослые самцы не обязательно более успешны при охоте группами, чем поодиночке. Гепарды иногда получают травмы во время охоты, хотя смертельные увечья случаются редко. В очень редких случаях гепарды могут есть падаль или возвращаться к брошенной добыче.

Гепарды охотятся визуально и в основном днем, особенно с 07:00 до 10:00 и с 16:00 до 19:00, в часы, когда конкуренты менее активны. Хотя ночная охота в Серенгети также регистрировалась. Используются пять основных методов охоты: 1) сидеть в засаде при приближении ничего не подозревающего стада; 2) медленно подойти к стаду на виду и перейти в спринт с расстояния 60-70 м; 3) начать рывок на расстоянии 600 м, если добыча ничего его замечает или новорожденное копытное кормится; 4) приближаться к бдительной добыче полупригнувшись, замирать на полпути или приседать когда добыча смотрит вверх и чередовать это поведение с рысью или неподвижным сидением до тех пор, пока не приблизится достаточно близко, чтобы начать атаку; 5) вспугнуть добычу из растительности и преследовать ее, как только она побежит. На равнинах Серенгети два последних упомянутых метода являются основными способами поимки добычи. Гепардам может потребоваться более 2 лет, чтобы освоить некоторые из этих методов охоты, в молодом возрасте в основном полагаясь последний метод.

Добычу гепард преследует на короткой дистанции. Более мелкая добыча сбивается быстрым ударом передней лапы. Добычу среднего и крупного размера обычно заваливает, цепляя ее когтем и потянув назад; более крупную добычу валит, забравшись ей на бедро и затем опрокинув. Крупную добычу душит, зажимая челюстями горло, на что требуется время. Более мелкую добычу либо хватают за морду, либо убивают укусом в череп или заднюю часть шеи. Добычу либо съедает на месте убийства, либо тащит в ближайшее укрытие. Гепардам обычно требуется время, чтобы прийти в себя после погони, иногда до целого часа. Едят они быстро, обычно начиная с брюшной поверхности, затем съедая печень и сердце.

Гепарды практически не пытаются защитить свою добычу от более крупных хищников и клептопаразитов, таких как львы (Panthera leo) и пятнистые гиены (Crocuta crocuta), в НП Серенгети могут терять до 13% своей добычи из-за клептопаразитов. Как следствие, гепарды избегают охоты в районах присутствия этих хищников. Зарегистрированы случаи, когда гепардов от добычи отгоняли леопарды (Panthera pardus), гиеновидные собаки (Lycaon pictus), бурые гиены (Hyaena brunnea), полосатые гиены (Hyaena hyaena), бабуины (Papio spp.) и даже стервятники.

Социальное поведение различное у полов и уникальное для кошачьих. Самки живут одни, если у них нет котят, самцы могут всю жизнь жить небольшими постоянными коалициями из 2-3 (реже 4) особей, тогда как остальные живут поодиночке. Большинство этих коалиций полностью состоят из однопометников, но примерно 30% коалиций Серенгети включают неродственного самца. Самец или группа самцов поддерживают непосредственную близость к одиночной самке или матери с детенышами, когда у нее той начинается течка. Такая ассоциация может длиться до 2 дней (хотя имелись наблюдения, как самцы общаются с самками без эструса и их детенышами в течение до 18 дней), и такие ассоциации могут объяснять наблюдения более крупные зарегистрированные размеры групп.

Самцы мало сотрудничают при ловле добычи; обычно один просто начинает охоту, а остальные следуют за ним. Коалиции, однако, предпочитают охотиться на более крупную добычу (например, взрослых антилоп гну), чем на одиночных мелких особей (например, газелей Томсона), но даже после раздела туши между двумя или тремя особями каждый в коалиции получает больше пищи, одиночный самец. Однако коалиции имеют более высокую выживаемость, чем одиночки, только тогда, когда имеется много других коалиций. Это указывает на тот факт, что коалиции получают выгоду от своих улучшенных конкурентных способностей в борьбе с другими самцами.
В Серенгети 43% пометов не от одного самца-отца, и не только от членов одной и той же коалиции самцов. Спаривание происходит быстро, длится менее минуты, и в дикой природе отмечалось очень мало. В неволе члены коалиции часто спариваются последовательно. Предполагая, что члены коалиции делят спаривающихся поровну, грубые расчеты показывают, что самцы распределяются в соответствии с размером группы и территориальным статусом таким образом, что каждый встречает эквивалентное количество самок на равнинах Серенгети. Существуют веские причины полагать, что одиночные самцы-кочевники находятся в невыгодном положении по сравнению с владельцами участков и до сих пор неясно, почему некоторые самцы остаются одиночками и кочуют на протяжении всей своей жизни.

В открытых равнинах северного Серенгети самки имеют огромные участки площадью более 800 км², следуя за ежегодными перемещениями газелей Томсона. С другой стороны, взрослые самцы демонстрируют две различные поведенческие линии. Территориальные самцы (резиденты) имеют небольшие участки, которые постоянно метят мочой, тогда как самцы-нерезиденты (бродячие) ходят по большей части равнин Серенгети и редко делают метки мочой. Нерезиденты более осторожны, чем резиденты, поскольку чаще находятся начеку; у них наблюдаются признаки физиологического стресса и увеличение заболеваемостей. Участки самцов-резидентов составляют в среднем 37 км², тогда как у нерезидентов они составляют в среднем 777 км² и перекрывают участки других. Самцы не занимают территории постоянно. Все они начинают кочевать, но некоторые остаются нерезидентами всю свою жизнь, другие становятся территориальными, а третьи, впервые встретившиеся как оседлые, впоследствии становятся кочевниками.

Беременность 90-95 дней. Несмотря на то, что спаривание не носит сезонного характера, роды наиболее часто регистрируются в январе-августе, хотя в Серенгети пик рождаемости отмечается в сезон дождей (ноябрь-май). Детёныши рождаются в укрытии среди высокой травы или густой растительности, в подлеске или среди скал. Самки поедают послед. В помете рождается до 8 котят, хотя обычно из логова в дикой природе (в Серенгети) выходит максимум 6, а до независимости доживает в среднем лишь 2. Соотношение полов не отличается от 1 : 1 при рождении. В Серенгети НП детеныши при рождении весят 350 г. Примерно до трех месяцев котята имеют уникальный покров из длинных голубовато-серых или дымчатых волос длиной 70–80 мм, скрывающих крошечные пятна на нижней части шерсти. Глаза открываются на 10–12 день, а полный комплект молочных зубов прорезывается примерно к 6 неделям; постоянные зубы формируются примерно к 8–9 месяцам. Детёныши в НП Серенгети растут в логове в среднем на 45 г/день, кормление молоком заканчивается примерно в 4 месяца.

Матери возвращаются в логово, чтобы выкармливать детенышей в течение 6–8 недель, в течение этого времени они регулярно переносят детенышей в новые логова, прежде чем детеныши начнут сопровождать свою мать на охоте с возраста 3–4 месяцев. Самки ловят пищу для своего потомства, иногда оставляя добычу живой и выпуская ее, позволяя детенышам преследовать и ловить добычу самостоятельно, обучая охотничьим навыкам. Во время развития детеныши активно участвуют в играх, которые в среднем занимают 3,4% времени в сутки. Хотя игривые детеныши иногда портят охоту своей матери, общее влияние на успех охоты считается минимальным.
Покинув мать, однопометники остаются группой братьев и сестер в основном в пределах своего натального участка в течение примерно шести месяцев, но самки отделяются от группы во время первой течки. Самцы расходятся за пределы своего натального участка, что помогает избежать инбридинга, в то время как самки более филопатричны, их участки в Серенгети перекрывают участки матерей на целых 60%.

Некоторые данные о демографии и репродуктивности гепардов, живущих на равнинах Серенгети, Танзания, были получены в течение 25-летнего периода (1969 по 1994 г.). Средний возраст достижения независимости 17,1 месяцев, самки рожают первый раз примерно в 2,4 лет, интервал между рождениями 20,1 месяцев, а средний размер помета, доживающего до независимости, 2,1 котенка. До момента обретения независимости в Танзании самки вырастили в среднем 1,7 детенышей, а средняя репродуктивная способность составила 0,36 детенышей в год, или 0,17 помета в год. В Серенгети средняя годовая смертность самок и самцов составляет соответственно 0,32 и 0,61 для детенышей в возрасте 1–2 лет; и 0,15 и 0,31 для взрослых. Смертность детенышей может быть чрезвычайно высокой. Согласно одному из исследований, в Серенгети 95% детенышей погибли до обретения независимости, в основном из-за хищников. Большая часть смертей произошла в первые несколько месяцев, и матери смогли быстро зачать после потери детенышей. Детоубийство, т. е. убийство детенышей чужими самками или самцами гепардов у этого вида не отмечено.

Некоторые матрилинии в НП Серенгети гораздо более успешны, чем другие, при этом 5 линий составляют 45% популяции местных гепардов. Почему репродуктивно успешные матери производят репродуктивно успешных дочерей и внучек, неизвестно, но есть предположение, что более осторожные самки имеют более высокий репродуктивный успех. Поскольку хищничество львов очень важно для репродуктивного успеха этого вида, увеличивается вероятность того, что дочери унаследуют некоторые аспекты осторожности своих матерей или тактики борьбы с хищниками.
Данные о продолжительности жизни разнятся. Самки Серенгети продолжают размножаться до 11,8 лет и могут прожить до 13,6 лет в дикой природе. Однако это максимальные сроки. Средняя же продолжительность жизни самок, которые остались с матерями 6,2 года. Самцы Серенгети живут максимум 7,8 лет, что намного меньше, чем самки. Минимальная продолжительность жизни самцов 2,8 года для тех, кто родился в исследуемом районе, но 5,3 года для иммигрантов.
В неволе максимальная продолжительность жизни гепарда составляет 21 год и большое количество животных проживает дольше 17 лет.

Важные охраняемые территории, которые представляют собой оплот популяций гепардов в Восточной Африке, включают НП Цаво и НП Руаха (Танзания) и, конечно же, заповедник Серенгети и экосистему Мара (Танзания, Кения).
Охота запрещена в Кении, Танзании и Уганде. Данных по Сомали нет.
Как и другим гепардам, восточно-африканскому угрожает браконьерство, потеря мест обитания, хищничество львов, леопардов, гиен и гиеновидных собак, что в первую очередь приводит к высокой смертности из детенышей. В экосистеме Серенгети смертность котят достигает 90-95%. Есть подозрение, что в первую очередь источниками этого являются масайские львы.
Гепарды особенно уязвимы к утрате и фрагментации среды обитания. Гепардам, живущим за пределами охраняемых территорий, часто угрожают конфликты с животноводами и фермерами.
Высокоскоростные дороги также представляют растущую угрозу для популяций гепардов. Это вызывает особую озабоченность там, где асфальтированные дороги пересекают или примыкают к основным территориям охраняемой дикой природы, например, дорога Найроби-Момбаса, которая пересекает НП Цаво в Кении. В период с 2014 по 2019 г. 6 взрослых гепардов были сбиты автомобилями на грунтовой главной дороге через НП Серенгети в Танзании.

Хотя гепард может поражаться инфекционными заболеваниями, особенно чесоткой в экосистеме Серенгети-Мара, низкая плотность гепарда делает маловероятным то, что инфекционные заболевания представляют серьезную угрозу для свободно перемещающихся популяций гепардов. В Северном Серенгети у гепардов наблюдается высокая серологическая распространенность кошачьего коронавируса и герпесвируса. Вирус иммунодефицита кошек также присутствует с высокой частотой (40%) в некоторых популяциях гепардов в Танзании.
В некоторых районах на гепардов охотятся из-за их шкур, а также в культурных целях. Кроме того, существует значительная незаконная торговля детенышами гепарда в качестве домашних животных в государства Персидского залива.



Литература
1. Krausman P. R., Morales S. M. "Acinonyx jubatus" / Mammalian Species. No. 771, pp. 1-6. Published 15 July 2005 by The American Society of Mammalogists.
2. Heller E. "Acinonyx jubatus raineyi, new subspecies. Rainey African Cheetah" / New Races of Carnivores and Baboons from Equatorial Africa and Abyssinia. Smithsonian Miscellaneous Collections, 1913. Volume 61. Number 19. pp. 9-10.
3. Heller E., Roosevelt T. "Rainey African Cheetah. Acinonyx jubatus raineyi" / Life-histories of African game animals. Volume I. New York, Charles Scribners's Sons, 1914. pp. 248-249.
4. Heller E. "Acinonyx jubatus velox, new subspecies. African Highland Cheetah" / New Races of Carnivores and Baboons from Equatorial Africa and Abyssinia. Smithsonian Miscellaneous Collections, 1913. Volume 61. Number 19. pp. 7-9.
5. Hilzheimer, M. "Über neue Gepparden nebst Bemerkungen über die Nomenklatur dieser Tiere". Sitzungsbericht der Gesellschaft naturforschender Freunde zu Berlin, 1913, no. 5, pp. 289-290.
6. Harper F. "East African Cheetah (Acinonyx jubatus ngorongorensis)" / Extinct and vanishing mammals of the Old World. Special publication No. 12, New York, 1945. pp. 278-280.
7. Kitchener A. C., Breitenmoser-Würsten Ch., Eizirik E., Gentry A., Werdelin L., Wilting A., Yamaguchi N., Abramov A. V., Christiansen P., Driscoll C., Duckworth J. W., Johnson W., Luo S.-J., Meijaard E., O’Donoghue P., Sanderson J., Seymour K., Bruford M., Groves C., Hoffmann M., Nowell K., Timmons Z., Tobe S. "A revised taxonomy of the Felidae. The final report of the Cat Classification Task Force of the IUCN/SSC Cat Specialist Group" / Cat News Special Issue 11, 2017. - 80 pp.
8. Prost S., Machado A. P., Zumbroich J., Preier L., Mahtani-Williams S., Guschanski K., Brealey J. C., Fernandes C., Vercammen P., Godsall-Bottriell L., Bottriell P., Dalton D. L., Kotze A., Burger P. A. "Conservation Genomic Analyses of African and Asiatic Cheetahs (Acinonyx jubatus) Across Their Current and Historic Species Range" / Molecular Ecology, 2022 Aug; 3(16): 4208-4223.
9. Caro T. "Acinonyx jubatus. Cheetah" / in Kingdon J. & Hoffmann M. (eds) "Mammals of Africa. Volume V. Carnivores, Pangolins, Equids and Rhinoceroses". Bloomsbury Publishing, London, 2013. pp. 187-196.
10. Nowel K., Jackson P. "Cheetah, Acinonyx jubatus, Schreber, 1776" / Wild Cats: Status Surway and Conservation Action Plan. IUCN/SSC Cat Specialist Group. Gland, Switzerland, 1996. pp. 12-16.
11. Durant S. M., Groom R., Ipavec A., Mitchell N., Khalatbari L. "Acinonyx jubatus" / The IUCN Red List of Threatened Species 2022: e.T219A124366642.
12. Khalatbari L., Durant S., Kitchener A.C., Breitenmoser-Würsten C., Ostrowski S., Brouwer E., Breitenmoser U. "The situation of the Cheetah in Asia". CMS, Bonn, Germany, 2022. - 136 pp.